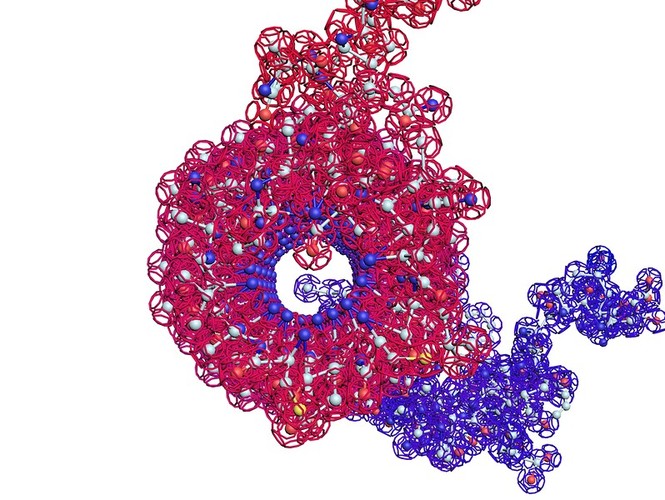
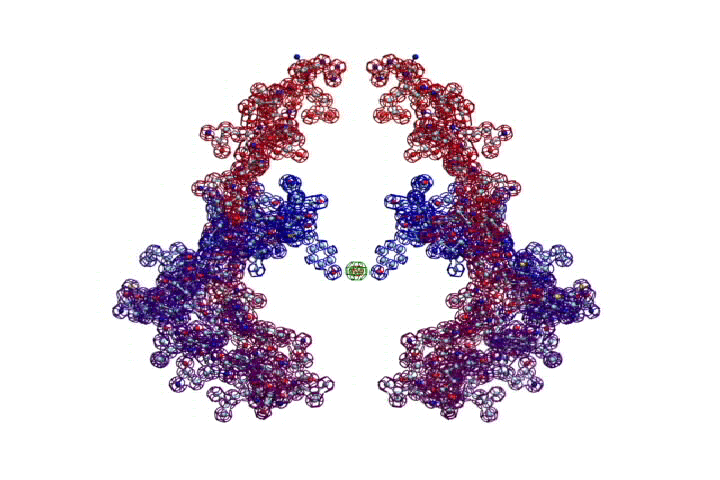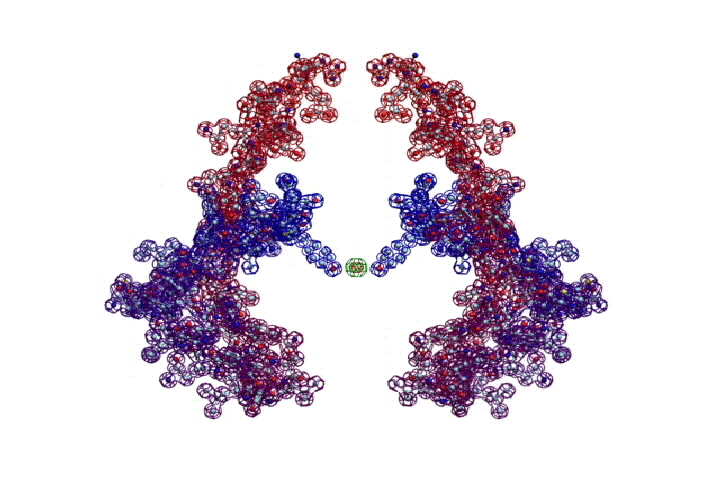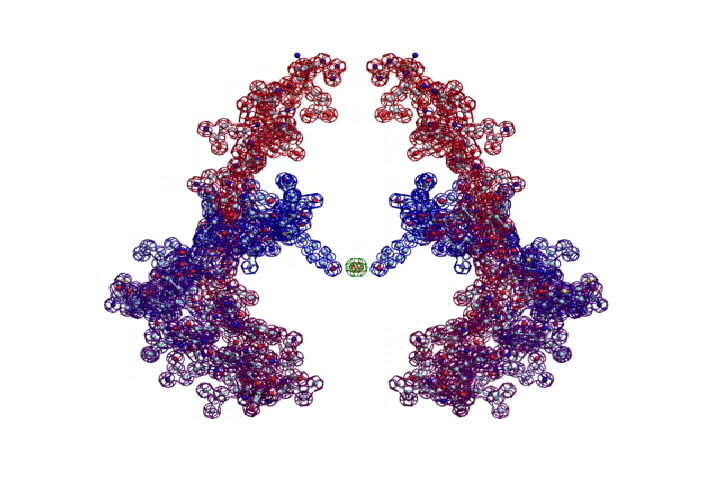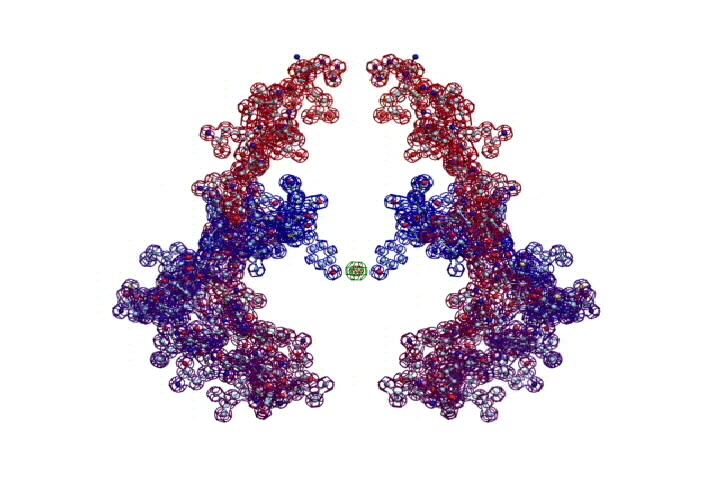
Повороты (изломы) спиралей на пролине
Композиционный, транспозиционный и пролиновый углы
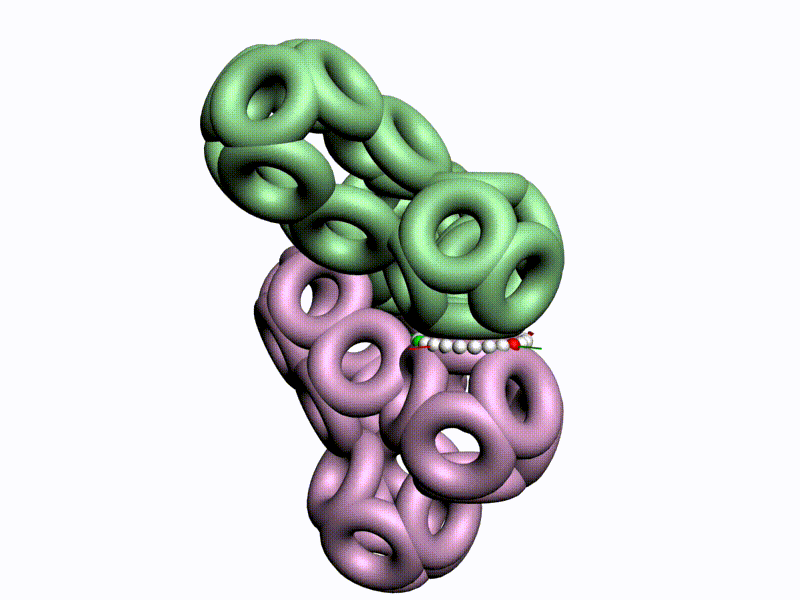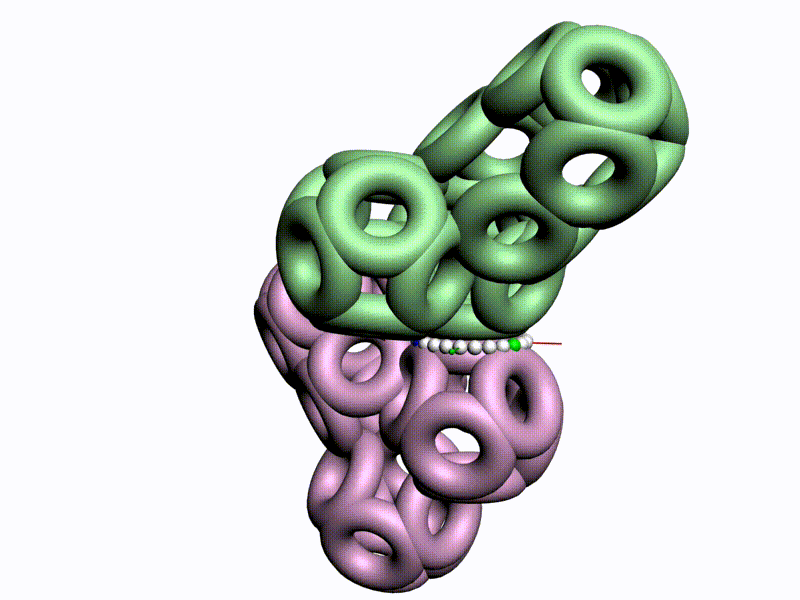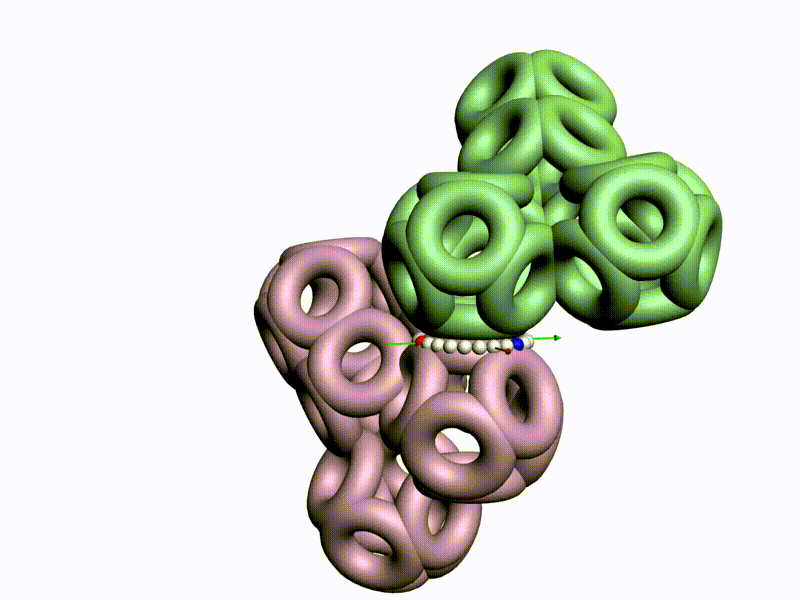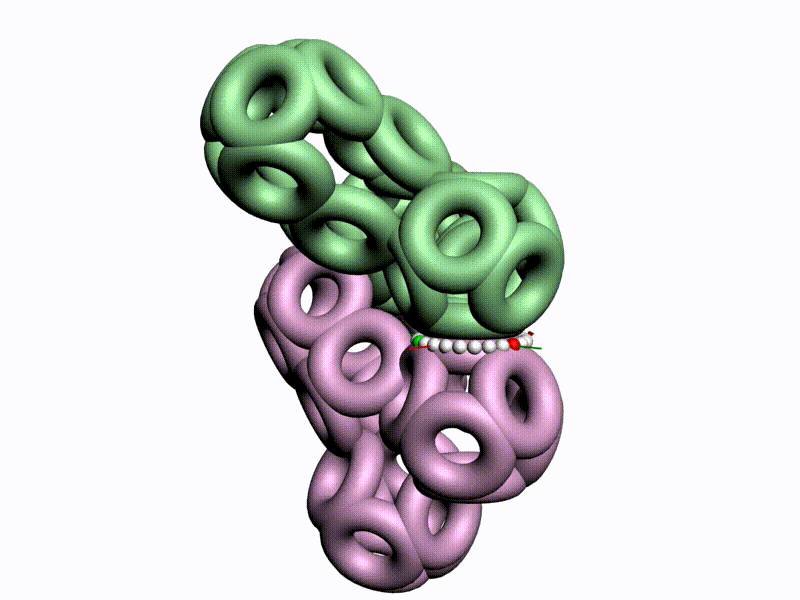
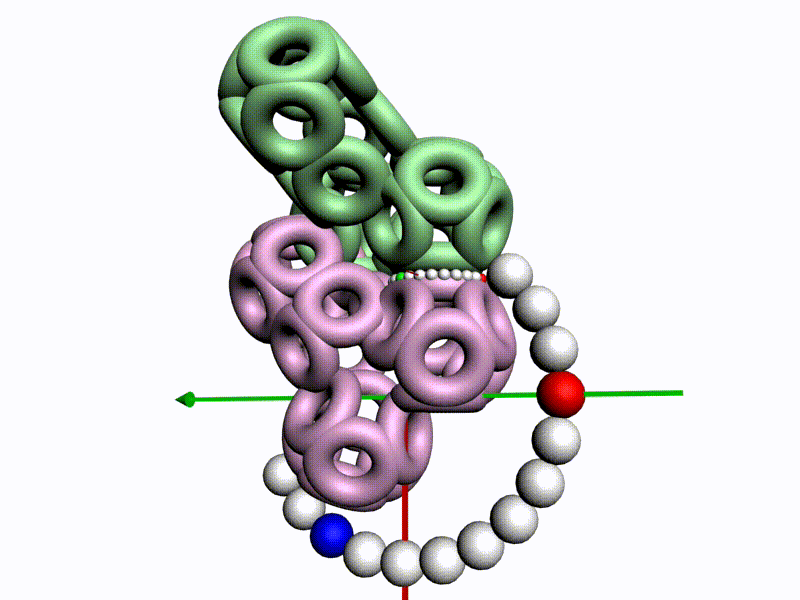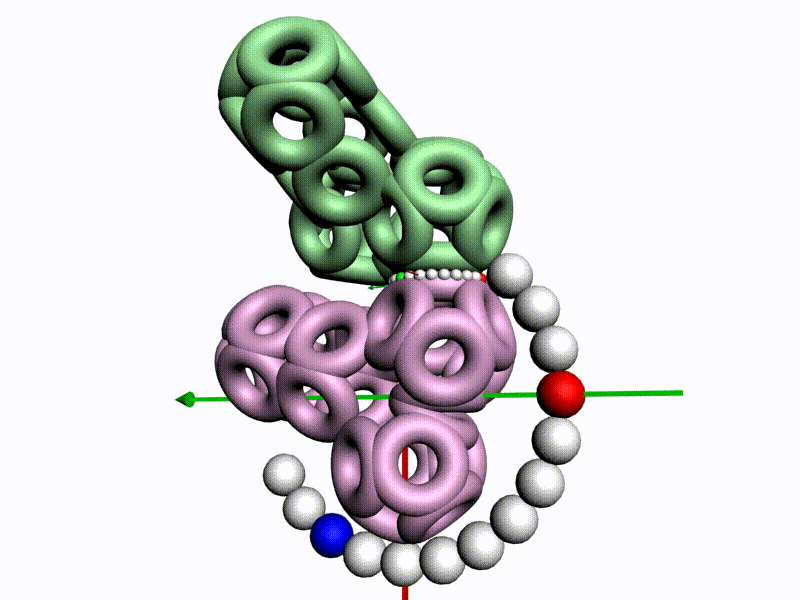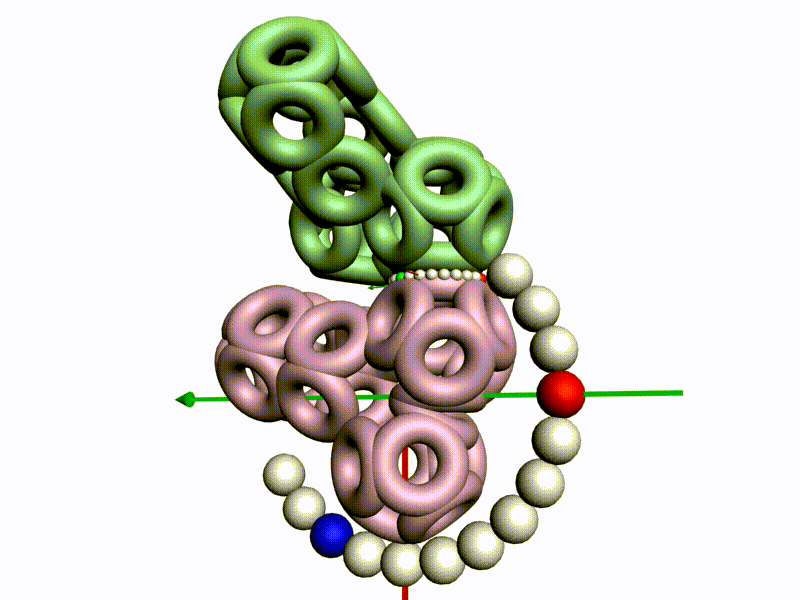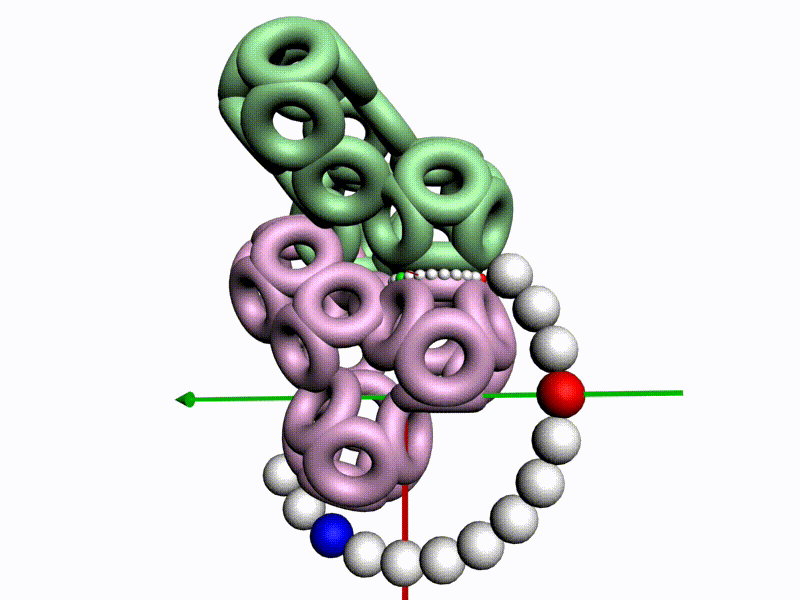
По шкале, изображенной в виде фрактального эталона, меняется значение композиционного угла.
Транспозиционный угол откладывается по шкале, которая лежит в перпендикулярной плоскости к шкале композиционных углов
Диапазоны и взаимная ориентация шкал изменения композиционного и транспозиционного углов показывают, что сборка белковой молекулы может идти почти по полному телесному углу на каждом шаге. Библиотеку готовых 3D конструкций можно взять из геномов разных организмов.
Красная отметка (принадлежит кольцевому электрону, центр которого находится в плоскости текущей пептидной группы) совмещается с кольцевым электроном предыдущего аминокислотного остатка, центр которого находится в плоскости симметрии предыдущей пептидной группы.
После того, как очередной аминокислотных остаток добавился в модель белка, модель продвинулась, повернулась и далее поворачивается уже на транспозиционный угол, значение которого меняется по шкале из более крупных шариков.
Транспозиционный угол фиксирован в альфа-, 310- и пи-спирали, но в каждой из них принимает своё значение. В бета-спирали он может меняться, если нет периодических водородных связей между соседними бета-спиралями, образующими бета-слой. В программных спиралях он периодически меняется, т.е. достаточно определить транспозиционные углы только для одного периода программной спирали.