
Модель антикодоновой петли тРНК



Метилинозин срезает продольную диэфирную связь, оставляя триплет иРНК на антикодоновой петле тРНК. тРНК продолжает по инерции вращаться, пока третий нуклеотид кодона иРНК не встретит комплементарное основание на сайте рибосомы.
Третий нуклеотид триплета терминирует угол поворота тРНК в рибосоме. После этого аминокислота устанавливается в растующую бековую молекулу под нужным углом.
Модель метилинозина оказалась ключевой при моделировании антикодоновой петли тРНК.
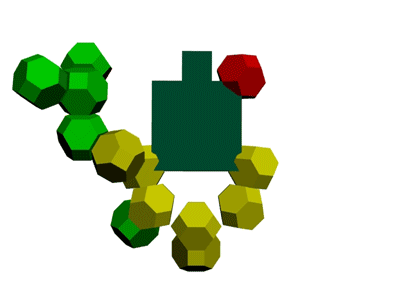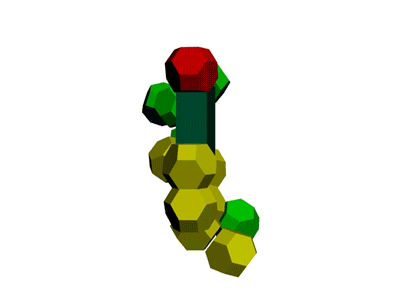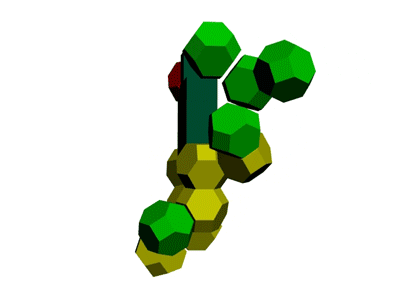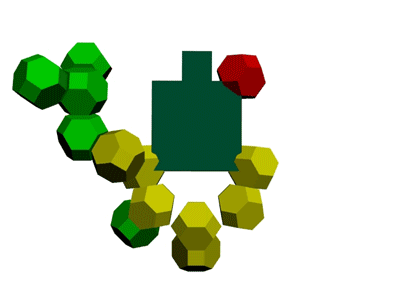
Образование водородной связи в канале рибосомы
В канале рибосомы присутствуют протоны.
Через миллисекунду после установки очередного аминокислотного остатка они образуют водородную связь -
ещё до того, как тРНК отпустит аминокислотный остаток, который она держит 4-мя вандерваальсовыми связями

Для каждой аминокислоты (в разных спиралях белка) эта ось (ось вращения tRNA) наклонена по-своему.
Для 310-спирали и альфа-спирали угол поворота tRNA отличается на 120-100=20 градусов. Среднее значение примем за начало отсчета, т.е. 0 градусов. Пи-спираль строится, если аминокислота повернута на 120 градусов от начала отсчета (от альфа-310), а бета-спираль строится, если аминокислота повернута на 240. Время установки аминокислот отличается. Для альфа-спирали оно минимально, для пи-спирали раза в два больше, а для бета-спирали еще раза в два больше. Поэтому сборка бета-спирали идет примерно в 4 раза медленнее, чем сборка альфа-спирали.
Кстати, угол поворота тРНК не совпадает с углом между остатками аминокислот в альфа-, 310-, пи-, бета-спиралях. Поэтому углы поворота tRNA отличаются на 120 градусов, а углы в белковых спиралях, т.е. уже вокруг оси симметрии белковых спиралей получаются ~120 для 310-спирали, ~100 для альфа-спирали, ~87 для пи-спирали и больше 180 для бета спирали. Это - чистая стереометрия.
Кушелев: В инернте для пи-спирали встречается два числа. 4.1 остатка на виток и 4.4. Какое из значений правильное?
число 3 для 310-спирали тоже нужно уточнить. Но это можно сделать только на достаточно длинных 310-спиралях. А официально считается, что длинных 310-спиралей не бывает. Это совсем недавно мне удалось обнаружить 310-спираль длиной более 1000 витков. Учитывая, что спирали белков упругие, понятно, почему для альфа-спирали пишут значение 3.6 ... 3.7 аминокислотных остатка на виток. Хотя в последнее время уже упростили до 3.6. Понятно, что и для 310-спирали на самом деле есть некий интервал типа 2.95 ... 3.05 аминокислотных остатка на виток. Это же относится и к пи-спиралям, и к бета-спиралям.
Итак, углы 0, а точнее -10 для альфа-спирали и +10 для 310-спирали, 120 для пи-спирали и 240 для бета-спирали соответствуют совсем другим углам, если их отсчитывать вокруг оси симметрии белковых спиралей.
Для 310-спирали это получается примерно 120 градусов, для альфа-спирали примерно 100 градусов, где-то я встречал значения 97...100 градусов, для пи-спирали 87 градусов, что соответствует 4.1 ... 4.2 остатка на виток. Для бета-спирали официально считается 180 градусов, что тоже нужно уточнять... На самом деле после настройки углов 310-, альфа- и пи-спиралей угол бета-спирали "лежит на поверхности". В бета-спирали на виток получается где-то 1.5 АО, а угол относительно оси спирали 240 градусов соответственно.
Выяснилось, что правильные углы между спиралями разных типов получаются при правильном задании начала отсчета углов. Другими словами, нужно поставить аминокислоту в правильную позицию, правильно сориентировать по всем трем осям, после чего получаются корректные модели белков. Но это только геометрический уровень. Нужно ещё учесть транспозиционные углы, которые отличаются в разных спиралях и зафиксированы водородными связями. Met и Pro - отдельный вопрос.
Поворот аминокислот в рибосоме и при вращении белковых спиралей происходит вокруг разных осей. Поэтому набору углов 0, 120, 240, которые отсчитываются вокруг оси вращения тРНК соответствуют углы 97..100, 119...121, ~87 и ~240 в альфа-, 310-, пи- и бета-спиралях.
Чтобы это понять, нужно посмотреть на тетраэдр сверху. Кажется, что между его гранями углы в 120 градусов. Но на самом деле в пространстве эти углы по 109 градусов 28 минут. Почему? А потому что они отмеряются не вокруг вертикальной оси, вдоль которой мы смотрим на тетраэдр сверху, а вокруг оси двухгранного угла, совпадающей с наклонным ребром тетраэдра.
Перечитываем книгу "Пикотехнология белков"...

Что мы видим на этой схеме? Поворот аминокислот вокруг оси tRNA на 120 градусов. Но оси симметрии белковых спиралей НЕ СОВПАДАЮТ с осью, вокруг которой поворачиваются аминокислоты! Этот факт не лежит на поверхности, но ... именно он превращает значения одних углов (0, 120, 240) в значения ДРУГИХ углов (альфа=97...100, пи=-86...88, бета). Если учесть это преобразование углов, то вместо неправильных 0, 120, 240 получатся правильные углы альфа-, бета-, пи-спиралей, которые не деформированы и не зафиксированы водородными связями. Если же учесть водородные связи, то альфа-угол будет изменен в альфа-спирали в одну сторону, а в 310-спирали в другую примерно на 10 градусов. Изменен он будет и в пи-спирали. Тоже на несколько градусов. Изменение коснётся не только композиционных, но и транспозиционных углов. Если это учесть, то геометрический алгоритм поможет получить более точную 3D модель. Учесть нужно и изменение в первую очередь транспозиционного угла в случае Met. Учет изменение третьего угла (угла Pro) даст дополнительную гибкость модели на остатках Pro. Это всё - чистая геометрия. Далее начнётся учет физико-химических взаимодействий. Но делать это нужно с учетом кольцевой формы электрона и соответственно кольцегранной формы аминокислотных остатков.

Аминокислота поворачивается вокруг вертикальной оси (оси симметрии tRNA)

Поворот на 120 градусов относительно оси симметрии tRNA формирует углы альфа-310-, бета-, пи-спиралей соответственно.
Акцепотрный стебель ориентирован вдолб оси вращения тРНК

Но это возможно лишь в том случае, если молекула тРНК симметрична относительно оси симметрии акцепторного стебля:

Это - модель ~половины тРНК, т.е. только одна цепь. Должны быть ещё обратные цепи, которые пока не показаны. Конечно, это ещё неправильная модель, но уже "близко к тексту", т.е. в первом приближении тРНК имеет именно такую форму.

Линейная модель ДНК/РНК
Пикотехнологический ДНК-роцессор
Все азотистые основания направлены вдоль оси симметрии ДНК. Протоны представляют собой "оперение", на которое натыкаются молекулы воды и подталкивают ДНК в одном направлении. ДНК начинает двигаться, а имея спиральную форму ещё и вращаться, что стабилизирует направление движения.

Упрощённая (многогранная) модель "Минор Кушелева". Особенностью минора Кушелева является ортогональность плоскостей азотистых оснований, образующих минорную комплементарную пару C-U / C-T
Пикотехнологический ДНК-процессор.
Даже в структуре суперспирали ДНК половина ОН-групп могут вращаться, т.е. ступени можно раздвигать. В двойной спирали ДНК вращаться могут 75% OH-групп. При этом не только можно раздвигать ступени и снимать копии, но и менять форму двойной спирали, например, формировать шпильки. В одинарной цепи ДНК/РНК вращаться могут 100% OH-групп. При этом цепь может наращиваться в произвольном направлении и фиксироваться. Для жёсткой фиксации формы используются комплементарные архитектурные перевертыши. Фосфатные группы ДНК/РНК по существу представляют собой пикотехнологический процессор, а точнее механизм, который меняет форму молекулы, которая может запоминаться за счёт взаимодействия комплементарных (и не очень) азотистых оснований. Удачная РНК-конструкция может тиражироваться, но может и модифицироваться, т.е. деградировать или развиваться. Эволюция, вероятно, началась или приобрела гигантский размах с разделения функций памяти и механизма, т.е. дифференцирования универсальных одноцепочечных ДНК на РНК и двухцепочечные ДНК.

Модель гексафосфата, который по существу является элементом "центрального" пикотехнологического процессора суперспирали ДНК. Число таких элементов в три раза меньше числа нуклеотидных пар, входящих в состав суперспирали ДНК.
Модель дигидроуридиловой петли
Моделирование центрального узла (перекрестия тРНК).
На схеме тРНК указаны два нуклеотида U-U, которые якобы образуют минорную пару. В действительности эти нуклеотиды образуют связи с другими цепями тРНК. Какими?
Это предстоит выяснить в предстоящем модельном эксперименте. Центральный узел тРНК наиболее сложный, поэтому его моделирование и отложено на последний этап.

Чтобы разобраться, как на самом деле расположены нуклеотиды на перекрестии тРНК, придётся собрать пластмассовую модель.
"Комплементарная" пара U-U (урацил-урацил) находится в исходной позиции, т.е. как показано на схеме в учебнике. Наша задача найти настоящие комплементарные пары для каждого урацила, а заодно смоделировать вариабельную петлю тРНК.
Пикотехнологическая модель дигидроуридиловой петли помогла сделать научное открытие!
Задача сделать пластмассовую модель перекрестия тРНК оказалась слишком сложной, поэтому было решено начать с более простой, а именно - с моделирования дигидроуридиловой петли, которая очень похожа на псевдоуридиловую и выполняет ту же функцию. Эти две петли похожи на правую и левую руки или крылья.
Дигидроуридиловая петля тоже оказалась сложнее, чем представлялось в начале.

Когда стало понятно, что невозможно изготовть модель дигидроуридиловой петли тем же способом, что и псевдоуридиловой петли, было решено начать с "руля", т.е. с нуклеотидов GC, которые в дигидроуридиловой петле выполняют ту же функцию, что и в псевдоуридиловой.
Была изготовлена модель, составленная из двух нуклеотидов в той же композиции, что и в псевдоуридиловой петле.
После этого мы замкнули петлю так же, как и псевдоуридиловую, т.е. нуклеотиды G и DHU образовали такую же комплементарную пару с поворотом на 180 градусов, как аденин и псевдоурацил в псевдоуридиловой петле.
Далее следует ещё одна пара G-DHU, а за ней несколько странная пара аденин-аденин (А-А), которая, конечно же не является комплементарной. Мы оставили разборки с А-А на потом и продолжили сборку петли, тем более, что её начало C-G и G-U у меня уже было собрано с самого начала.
И тут мы заметили, что G-U как бы начинает развигаться, т.е. образует лишь одну водородную связь между центральным выступающим атомом гуанина и боковым выступающим атомом урацила.
Пара А-А ещё больше "разъехалась". При этом оба аденина образовали водородные связи с фосфатными группами, т.е. проявилось А-минорное взаимодействие второго рода (первого рода - взаимодействие центрального выступающего атома с одним из углеродов рибозы). В данном случае боковые атомы аденина взаимодействуют с образованием водородной связи с ОН-группами фосфатных групп.
А следующая пара дигидроурацил-гуанин (DHU-G) по-настоящему комплементарны с переворотом на 180 градусов. Далее следует G-DHU и "руль" GC.
Оказалось, что двойная спираль РНК может плавно разворачиваться на 180 градусов, словно книжка. Для этого достаточно сделать последовательность нуклеотидов, как в дигидроуридиловой петле.
Почему дигидроуридиловая петля устроена несколько иначе, чем псевдоуридиловая? Дело в том, что левый "руль" тРНК должен быть ориентирован зеркально симметрично правому "рулю". А двойная спираль-то закручена в одну сторону... Если сделать "плечи" тРНК одинаковыми, то второй "руль" будет "смотреть" не вперёд, а назад, и движение тРНК будет непрямолинейным. Поэтому второе "плечо" тРНК имеет дополнительный угловой набег на 180 градусов практически при той же длине. Достигается это плавным раздвиганием двойной спирали. Зная такие тонкости программируемой архитектуры тРНК можно приступать к моделированию самого сложного узла, а именно - перекрестия тРНК.
Кушелев: Попробуйте найти противоречия в моей модели антикодоновой петли тРНК, где не только прямой участок, но и точное позиционирование "ножа"-метил-инозина, отрезающего триплет иРНК. Я уже объяснял, что случайностей такого рода не бывает. Их вероятность - триллионная доля.
Виктория Соколик: С точки зрения биологии, это не эффективно. На каждой молекуле мРНК обычно усаживается десяток рибосом с формированием комплекса полисомы, которые и транслируют одновременно несколько копий белка. А Вы рассказываете, что матрица кромсается "ножом"-метил-инозина каждой тРНК на каждом кодоне и при этом "не случайно".
Кушелев: Я уже думал над этим. С первого взгляда кажется, что есть противоречие, но мы прекрасно знаем, что существуют ферменты, обрезающие повторы ДНК при делении клеток, но есть и ферменты, восстанавливающие повторы ДНК (концы хромосом). Следовательно триплеты иРНК тоже могут и обрезаться, и восстанавливаться. С первого взгляда может показаться, что это бессмысленно, но если иначе не удаётся управлять углом поворота аминокислоты, то обрезание с последующим восстановлением вполне оправдано.
тРНК входит в рибосому и за счёт своей энергии движения срезает триплет тРНК. После этого она продолжает вращаться по инерции вместе с отрезанным триплетом и останавливается в том положении, где третий нуклеотид триплета попадает на комплементарный нуклеотид рибосомы. Далее рибосома устанавливает аминокислоту в нужной позиции, после чего отделяет триплет иРНК от антикодона тРНК и присоединяет его к концу иРНК, которая выходит из рибосомы. Снаружи кажется, что иРНК вошла и вышла из рибосомы без изменений, а на самом деле каждый триплет был отрезан, прокручен, отсоединён от тРНК и прикреплён к выходной части иРНК. Вполне реальный конвейер.
Третий закон термодинамики
Кушелев: Согласно третьему началу термодинамики (информационному закону Кушелева) информация, как и энергия, распределяется по степеням свободы равномерно. Это означает, что структура гиперакустических сигналов соответствует по сложности пространственной структуре макромолекул, в частности ДНК/РНК, белков и т.д.
Рибосома и тРНК возникли в добелковом мире, поэтому их сигналы проще, чем сигналы, предназначенные белковым пикомашинам. Возможно, что сигнал рибосомы синусоидальный или прерывистый. Но система навигации тРНК существует, т.к. обнаружены два резонансных "руля".
Виктория Соколик: Надеюсь, вы придёте к пониманию третичной структуры тРНК в виде буквы L. Я не теряю надежду smile.
Кушелев: По поводу буквы L я уже написал, что тРНК складываться по диагонали, превращаясь из креста в "букву L". Я даже предполагал, что после синтеза тРНК они хранятся в неактивной L-конформации, а по сигналу рибосомы активируется только нужная тРНК и раскладывается из L в +. Но теперь я считаю, что тРНК синтезируются и сразу переводятся в активное состояние. Далее идёт сортировка тРНК на рибосомах. Те, которые не подходят, "отфутболиваются", а подходящие отрезают триплет и отдают таким образом запасённую во время движения энергию рибосоме. При наращивании выходной части тРНК энергия возвращается рибосоме. Кстати, какое количество АТФ потребляет рибосома в пересчёте на один аминокислотный остаток?

Первый набросок пикотехнологической модели дигидроуридиловой петли тРНК. Углы ещё не отъюстированы, но форма в первом приближении такая. В принципе можно переходить к моделированию самого сложного узла тРНК, перекрестия с вариабельной петлёй.



Более точная пикотехнологическая модель дигидроуридиловой петли помогла сделать очередное научное открытие. Двойная спираль РНК может плавно раздвигаться на минорных комплементарных (и не очень) основаниях.

Пикотехнологическая модель псевдоуридиновой петли тРНК показывает, что расположенный на краю петли цитозин обладает подвижностью, является резонансной системой, которая обладает свойствами детектора акустических колебаний, источником которых является рибосома. Одна из двух (псевдоуридиновая или дигидроуридиновая) петель, которая находится ближе к источнику гиперзвука, возбуждается сильнее, среднее положение цитозина смещается, увеличивая длину петли. В результате, петля начинает двигаться медленнее, и тРНК заворачивает на источник гиперзвука, т.е. к рибосоме.
Схематическое изображение тРНК. На краях псевдоурациловой и дигидроурациловой петель изображены азотистые основания (цитозины) в невозбуждённом состоянии. В отсутствии акустического сигнала рибосомы тРНК движется по прямой, вращаясь вокруг большой оси.

Если же появляется акустический сигнал рибосомы, то цитозины начинают колебаться, причём амплитуда колебаний возрастает, если цитозин приближается к рибосоме. При этом ближний к рибосоме цитозин сильнее тормозит, в результате чего тРНК заворачивает к источнику акустического синала, т.е. к рибосоме.

Упрощённая модель тРНК, визуализирующая механизм навигации тРНК.
Крайние цитозины псевдоурациловой и дигидроурациловой петель тРНК подвижны и являются элементами резонансного рулевого управления тРНК. Амплитуда асимметричных колебаний цитозинов тем выше, чем ближе они к источнику гиперзвука, которым является активный центр рибосомы. Таким образом, тРНК плывут на зов рибосомы.
Итак, тРНК устанавливает по "быстрому" кодону очередную аминокислоту в растущую альфа-спираль, после чего начинает вращаться вместе с белком и матричной РНК на угол альфа-спирали, который примерно равен углу между соседними кодонами мРНК.
Однако тут есть загадка. Ведь аминокислота и антикодоновая петля находятся в разных местах. Как же они могут вращаться на один и тот же угол?
Это возможно в том случае, если ось альфа-спирали совпадает или почти совпадает с осью мРНК. Источник
Механизм трансляции
Сегодня исследователи механизма трансляции наивно полагают, что сначала белок отсоединяется от тРНК, а потом чудесным образом подсоединяется в аминокислоте, которую держит следующая тРНК. В действительности сначала тРНК вставляет аминокислоту в растующую белковую "цепь", после чего мРНК начинает проворачиваться на 108 градусов, увлекая за собой белок и обе тРНК. Предыдущая тРНК за что-то цепляется и отваливается от построенного участка белковой молекулы, т.к. энергия связи 4 вандерваальсовых связей меньше, чем энергия связи аминокислотного остатка спереди и сзади.
За поворотом следует реверс, поэтому повёрнутый белок не меняет взаимной ориентации двух тРНК.
Кушелев: Удаление предыдущей тРНК происходит при вращении мРНК в случае быстрого кодона и при реверсе новой тРНК в случае медленного кодона.
На данной анимации показан реверс тРНК вместе с белковой "цепью", в начале которого должна "отвалиться" предыдущая тРНК


Перевод новой тРНК в положение предыдущей происходит с помощью продвижения мРНК на один кодон. При этом предыдущая тРНК автоматически "отваливается", а белковая "цепь" поворачивается на угол альфа-спирали, т.е. на 97-100 градусов перпендикулярно оси симметрии новой тРНК.. Это меньше, чем угол 108 градусов, на который поворачивается мРНК, поэтому антикодон повёрнутой тРНК "отваливается и от кодона мРНК. Во время реверса новая тРНК "отламывает" старую тРНК и от растущей белковой "цепи", вращая белок по другой оси. Этот механизм возник в процессе эволюции композиционного генетического кода, т.е. с появлением универсальных тРНК. До этого каждому триплету соответствовала своя тРНК. В процессе эволюции число типов тРНК сократилось.
Цитата: меет типичную длину от 73 до 93 нуклеотидов и размеры около 5 нм. Конец цитаты.

тРНК (общепринятая модель)
РСА не позволяет определить форму тРНК. РСА не отличает крест от уголка
Дальше процесс строительства белковой "цепи" повторяется

Перевод новой тРНК в положение предыдущей происходит с помощью продвижения мРНК на один кодон. При этом предыдущая тРНК автоматически "отваливается", а белковая "цепь" поворачивается на угол альфа-спирали, т.е. на 97-100 градусов перпендикулярно оси симметрии новой тРНК.. Это меньше, чем угол 108 градусов, на который поворачивается мРНК, поэтому антикодон повёрнутой тРНК "отваливается и от кодона мРНК. Во время реверса новая тРНК "отламывает" старую тРНК и от растущей белковой "цепи", вращая белок по другой оси. Этот механизм возник в процессе эволюции композиционного генетического кода, т.е. с появлением универсальных тРНК. До этого каждому триплету соответствовала своя тРНК. В процессе эволюции число типов тРНК сократилось. Ссылка
Детализированная дискуссия
Пятый домен рибосомальной тРНК
Кольцегранная и многогранная (упрощённая) линейные модели ДНК / РНК.


Структурные формулы азотистых оснований




Виртуальныйпикотехнологичский ДНК-конструктор



ДНК построена на ортогональных геометрических элементах с пятиричной симметрией.
Упрощённая модель нуклеотида помогла мне сделать очередное научное открытие. Оказалось, что ДНК построена на ортогональных геометрических элементах с пятиричной симметрией. При этом заполнение пятиэлементных систем, которые являются элементами следующего фрактального уровня, неполное, поэтому система обладает свойством тонкой настройки параметров вблизи идеальной пятиричной симметрии.



Виктория Соколик: Виртуальная кольцегранная модель рибозы и других сложных органических молекул важна, нова и значима для узкого круга специалистов, которые с её помощью могут обосновать закономерности кодирования и трансляции генома, предусмотреть механизм создания новых синтетических белков с заданными свойствами и функциями и многое, многое другое. Для остальной части человечества данная модель настолько же значима в повседневной жизни, как для Вас, например, интеграл. Как часто Вы нуждаетесь в интеграле и какая Вам лично от него польза? А те ученые, которые изобрели для Вас мобильники, интернет, компьютер и т. д., вряд ли смогли бы обойтись без интеграла.
II Международная виртуальная конференция "Актуальные проблемы биохимии и бионанотехнологии" (15-18. 11.2011),Казань
ЗАГАДКА ИЗОАКЦЕПТОРНЫХ тРНК
В.В. Соколик
ГУ “Институт неврологии, психиатрии и наркологии АМН Украины”
В 1957 г. F.H.C. Crick выдвинул “адапторную гипотезу”, в которой пред-ложил на роль посредника между кодонами и аминокислотами в процессе матричного синтеза белка тРНК [1]. Далее было установлено, что количество различных видов тРНК в цитоплазме эукариот всегда больше, чем количество различных аминокислот и аминоацил-тРНК-синтетаз. Это значит, что не-сколько изоакцепторных тРНК могут быть узнаны одной и той же аминоа-цил-тРНК-синтетазой и соответственно нести на своём акцепторном стебле один и тот же аминокислотный остаток. Для большинства аминокислот изоак-цепторные тРНК имеют различные по первому нуклеотиду антикодоны и уз-нают различные по третьему нуклеотиду кодоны.
У прокариот и в органелах эукариот (митохондрии и хлоропласты) для каждой аминокислоты предусмотрен единственный вид тРНК, соответствующий всем кодонам данной аминокислоты. Поэтому мы можем сделать вывод, что изначально дуплетом первых двух нуклеотидов кодонов кодировалась лишь аминокислотная последовательность и необходимость в пуле изоакцепторных тРНК не наблюдалась. Для тРНК простеиших характерно отсутствие модификаций первого нуклеотида антикодона, что обусловливает возможность нестрогого соответствия каноническому взаимодействию нуклеотидов, а именно: первый нуклеотид антикодона тРНК “узнаёт” все четыре разновидности (A, G, U и C) нуклеотидов в третьей пози-ции кодонов. Более того, в структуре тРНК органелл и прокариот нередко от-мечается частичная редукция тимидил-псевдоуридиловой и дигидроуридило-вой петель.
Дальнейшая эволюция механизма трансляции шла по пути посттранс-крипционной модификации первого нуклеотида антикодона и совершенство-вания пространственной структуры тРНК. Химические модификации первого нуклеотида антикодона тРНК привели к формированию семейств изоакцеп-торных тРНК с ограничениями к формированию канонических пар с любым нуклеотидом в третьей позиции кодонов. В настоящее время идентифицирова-но несколько десятков модифицированных нуклеотидов в структуре тРНК, однако нас, прежде всего, интересуют выявленные модификации первого нук-леотида антикодона. Так, в цитоплазме эукариот редко встречаются тРНК с немодифицированным нуклеотидом в этой позиции: как правило,
• аденин (А) в результате окислительного дезаминирования всегда превра-щается в инозин (І);
• уридин (U) модифицируется до 5-метоксиуридина (mo5U), 5-карбоксиметоксиуридина (cmo5U), 5-(метиламинометил)-2-тиоуридинa (mnm5s2U), 5-(метоксикарбонилметил)-2-тиоуридинa (mcm5s2U), 5-(метиксикарбонилметил)уридинa (mcm5U);
• гуанин (G) и цитозин (С) иногда могут присутствовать в немодифициро-ванном виде, хотя чаще метилированы по 2'-гидроксилу рибозы (Gm и Cm);
• в некоторых тРНК в первом положении антикодона встречается гипермо-дифицированное производное гуанина – кьюозин (Q).
В данной работе мы постараемся ответить на вопрос: для чего у каждой аминокислоты появился пул изоакцепторных тРНК (вместо одной единствен-ной, как у прокариот) да ещё и с модифицированными нуклеотидами в первой позиции антикодона.
Анализ базы данных верифицированных нуклеотидных последова-тельностей тРНК (tRNAdb: http://trnadb.bioinf.uni-leipzig.de/) показал, что у эукариот аминокислоты, которые характеризуются полным семейством кодо-нов (все четыре разновидности нуклеотидов в третьей позиции), имеют тРНК, в первом положении антикодонов которых представлен инозин, способный узнавать С и G, но не А или U. Исключением являются Gly и Ile. ТРНКGly, не-смотря на четыре разновидности кодонов для этой аминокислоты, в первой позиции антикодона несут либо С, либо G. В первой позиции антикодонов тРНКIle присутствует І, хотя изолейцин детерминирован лишь тремя кодонами.
Модифицикация уридина в первой позиции тРНК ограничивает его спаривание с пиримидиновыми азотистыми основаниями, т.е. modU “узнаёт” только A или G. Q и его модификации характерны для первого нуклеотида антикодона тРНК тех аминокислот, которые кодируются только двумя видами кодонов с С или U в третьем положении (His, Asp, Asn, Tyr и другие). При этом для кодонов XYС предусмотрены тРНК с GYZ антикодоном. Поэтому надо полагать, что кьюозин специфично “узнаёт” уридин в третьей позиции кодонов данных аминокислот.
Обращает на себя внимание, что в пуле из 200 нуклеотидных последо-вательностей цитоплазматических тРНК эукариот (tRNAdb) 73% имеют в пер-вой позиции I, C или G, “узнающие” кодоны с C/G в третьем положении, 20% с U и modU для кодонов XYА вида и только 7% с Q для XYU кодонов. Объек-тивная предпосылка такого неравного распределения кроется в различной частоте встречаемости кодонов, которые комплементарны данным тРНК. Например, для генов Human, Mouse и Drosophila встречаемость кодонов с C/G, А или U в третьем положении соотносится как 74 : 13 : 13. Легко видеть, что XYC/G-кодоны аминокислот и их тРНК для спиральных мотивов белка составляют подавляющее большинство. Неудивительно, что проф. А.С. Спирин, отклоняясь от постулата о матричном синтезе белка в виде развёрнутой полипептидной цепочки, предположил, что в самой рибосоме полипептид синтезируется сразу в виде правой ?-спирали и по желобу выталкивается наружу по мере трансляции мРНК [2]. Такое обобщение хорошо в виде правила для большинства “спиральных” кодонов, исключением из которого является синтез фрагментов полипептидной цепи в виде ?-тяжа или витка левой 3/10 спирали с помощью тРНК, в первой позиции антикодона которых стоят нуклеотиды (U, N, Q), “узнающие” “не спиральные” кодоны.
В наших предыдущих публикациях была представлена гипотеза о том, что у эукариот сформировался механизм детерминирования в геноме структурного шаблона белка (а не только его аминокислотной последователь-ности), в декодировании которого при матричном синтезе непосредственное участие принимают различные виды изоакцепторных тРНК [3, 4]. В частности для аминокислот с полным набором кодонов эволюция предусмотрела три разновидности изоакцепторных тРНК (для остальных аминокислот две или одну):
• с инозином в первой позиции антикодона тРНК “узнающие” “спиральные” (XYС или XYG) кодоны;
• с уридином и его модификациями в первой позиции антикодона тРНК “узнающие” XYА кодоны;
• с кьюозином в первой позиции антикодона тРНК “узнающие” XYU кодоны.
Эти три типа изоакцепторных тРНК кроме первого нуклеотида антикодо-на различаются между собой пространственной структурой, точнее углом поворота акцепторного стебля вокруг своей оси (рис. 1). Последнее достига-ется спецификой взаимодействия тимидил-псевдоуридиловой и дигидроуридиловой петель в структуре тРНК. А именно: С в третьей позиции тимидил-псевдоуридиловой петли взаимодействует с G во втором положении дигидроуридиловой петли в структуре тРНК с U и modU в первой позиции антикодона, с G в четвёртом положении дигидроуридиловой петли в структуре тРНК с I, C или G в первой позиции антикодона и с G в шестом положении дигидроуридиловой петли в структуре тРНК с Q в первой позиции антикодона. Таким способом транслируется информация третьего нуклеотида кодона о конфигурации пептидной связи, которой аминокислота на АСС-конце тРНК будет присоединена к растущей полипептидной цепи в рибосоме. Очевидно, что тРНКAla с I в первой позиции антикодона присоединит аланин к растущему полипептиду пептидной связью в R-конфигурации, а трансляция четырёх и более подряд кодонов GСС/G приведёт к формированию витка правой ?-спирали полиаланина в желобе рибосомы. Аланин на АСС-хвосте тРНКAla с U в первой позиции антикодона (для кодонов GСА) будет присоединён пептидной связью в 0-конфигурации, что сформирует ?-тяж. Аланин в составе комплекса с тРНКAla, первый нуклеотид которой кьюозин, присоединится пептидной связью в L-конфигурации. Эти изоакцепторные тРНКAla “узнают” GСU кодоны аланина, а троекратное их повторение детерминирует виток левой 3/10 спирали. Очевидно, что чередование “спиральных” и “неспиральных” кодонов транслируется в виде неструктурированного фрагмента без элементов вторичной структуры в виде хаотично изогнутой полипептидной цепи.
Таким образом, изоакцепторные тРНК необходимы для трансляции закодированной в геноме информации о конфигурации пептидных связей в транслируемом структурном шаблоне белка, а не только аминокислотной по-следовательности, как у прокариот.
Литература
1. Crick F.H.C. The Origin of the Genetic Code // J. Mol. Biol. — 1968. — 38. — Р. 367-379.
2. Спирин А.С. Молекулярная биология: рибосомы и синтез белка. – М: «Академия», 2011. – 496 с.
3. Sokolik V.V. Protein is coded in genome and synthesized in ribosomes as a structural template of a rotameric version sequence of peptide bound configura-tion / The International Moscow Conference on Computational Molecular Biology, МССМВ-11 (July 21-24, 2011), Moscow, Russia. – P. 347-348.
4. Соколик В.В. Способ моделирования пространственной структуры белка по детерминирующей его нуклеотидной последовательности // Биофизиче-ский вестник (ХНУ). – 2010. - 24 (1). – С. 31-45.
213
Дополнения
1
Сводная таблица первых нуклеотидов антикодонов тРНК по надцарствам биообъектов, которые уже идентифицированы экспериментально:

http://fotki.yandex.ru/users/sokolik67- … ew/402705/
Голубым цветом в столбце эукариот обозначены нуклеотиды, которые распознают спиральные кодоны (т.е. при трансляции на рибосоме формируется R-конфигурация пептидной связи), желтым цветом -- нуклеотиды, распознающие неспиральные кодоны для бета-тяжей (0-конфигурация пептидной связи) и розовым -- нуклеотиды, распознающие достаточно редкие кодоны левой спирали (L-конфигурация пептидной связи).
2
Здесь можно познакомиться с перечнем известных модификаций нуклеотидов:




http://fotki.yandex.ru/users/sokolik67- … ew/402701/
http://fotki.yandex.ru/users/sokolik67- … ew/402702/
http://fotki.yandex.ru/users/sokolik67- … ew/402703/
http://fotki.yandex.ru/users/sokolik67- … ew/402704/
3
Модели некоторыъ канонических и неканонических пары нуклеотидов, которые должны формироваться между первым нуклеотидом антикодона тРНК и третьим нуклеотидом кодона мРНК:








http://fotki.yandex.ru/users/sokolik67- … ew/402707/
http://fotki.yandex.ru/users/sokolik67- … ew/402708/
http://fotki.yandex.ru/users/sokolik67- … ew/402709/
http://fotki.yandex.ru/users/sokolik67- … ew/402710/
http://fotki.yandex.ru/users/sokolik67- … ew/402711/
http://fotki.yandex.ru/users/sokolik67- … ew/402712/
http://fotki.yandex.ru/users/sokolik67- … ew/402713/
http://fotki.yandex.ru/users/sokolik67- … ew/402714/
4
Еще несколько возможных и невозможных взаимодействий нуклеотидов, которые не обязательно встречаются в узнающих друг друга триплетах, но значимы для формирования пространственной структуры тРНК:



http://fotki.yandex.ru/users/sokolik67- … ew/402715/
http://fotki.yandex.ru/users/sokolik67- … ew/402716/
http://fotki.yandex.ru/users/sokolik67- … ew/402717/
5
Результат такого моделирования на основе структурного соответствия азотистых оснований обобщен в таблице:

http://fotki.yandex.ru/users/sokolik67- … ew/402706/
где косые линии между парами нуклеотидов обозначают отсутствие взаимодействия, а горизонтальные -- количество водородных связей между азотистыми основаниями нуклеотидов.
Девиация угла наклона ступеней ДНК/РНК
Кушелев: Методом "реверсной кинематики", а по-простому - автоматизированного подгона, удалось определить углы наклона ступеней ДНК более точно. Они оказались больше 70, но меньше 90 градусов. Вероятно, реальный наклон ступеней ~81...82 градуса, и этот наклон может меняться в широком интервале, т.е. от ~70 до ~90 градусов.

Вараллельно с возникновением водородной связи между азотистым основанием аденина (А) и третьей CH2-группой рибозы другого нуклеотида возникает взаимодействие между фосфатными группами этих же нуклеотидов. Взаимное расположение фосфатных групп напоминает взаимное расположение радикалов лейцина в лейциновых "молниях-застёжках" белков.
Для А-минорного взаимодействия характерно, что "А-стопки" в действительности представляют собой классическую одноцепочечную спираль РНК, ось симметрии которой совпадает с осью симметрии двухцепочечной спирали, с которой взаимодействует одноцепочечная. Таким образом, длинные участки с А-минорным взаимодействием и трёхцепочечная (или трёхнитевая) РНК - это одна и та же структура
Маленьким белым кубиком изображена водородная связь между аденином и второй(?) группой CH2 рибозы другого нуклеотида. Фосфатные группы аденина показаны голубым цветом, чтобы легче было отличить от фосфатных групп других нуклеотидов.


Проверяем модель трансляции!
Итак, средняя кинетическая энергия молекулы (10^-20 Дж) на порядок меньше энергии диэфирной связи (10^-19 Дж), которую могла бы отрезать тРНК за счёт своей кинетической энергии. Однако нужно учесть, что энергия молекулы тРНК существенно превосходит энергию молекулы воды, оценку которой мы получили. Дело в том, что тРНК накапливает кинетическую энергию при взаимодействиях с молекулами воды. Допустим, что скорость движения тРНК в три раза меньше скорости движения молекул воды. При этом энергия каждого атом типа углерода, который входит в состав тРНК будет на порядок меньше, т.е. 10^-21 Дж. Но молекула тРНК состоит, например, из 76 нуклеотидов, каждый из которых состоит из десятков атомов. Поэтому кинетическая энергия тРНК на три порядка больше кинетической энергии молекулы воды, т.е. 10^-18 дЖ, что на порядок превышает энергию диэфирной связи. Отсюда следует, что гипотеза отрезания триплета иРНК за счёт кинетической энергии тРНК в рибосоме подтверждается...

Модель аланина ( Ala/A по коду gcc )

Модель аргинина-top ( Arg_top/R по коду cgc )

Модель аргинина-bottom ( Arg_bottom/R по коду agg )

Модель аспарагина ( Asn/N по коду aac )

Модель аспарагиновой кислоты ( Asp/D по коду gac )

Модель цистеина ( Cys/C по коду tgc )

Модель Глутамина ( Gln/Q по коду cag )

Модель глутаминовой кислоты ( Glu/E по коду gag )

Модель глицина ( Gly/G по коду ggc )

Модель гистидина ( His/H по коду cac )

Модель изолейцина ( Ile/I по коду atc )

Модель лейцина-lamda ( Leu_lamda/L по коду ctc )

Модель лейцина-teta ( Leu_teta/L по коду ttg )

Модель лизина ( Lys/K по коду aag )

Модель метионина ( Met/M по коду atg )

Модель фенилаланина ( Phe/F по коду ttc )

Модель серина-teta ( Ser_teta/S по коду agc )

Модель треонина ( Thr/T по коду acc )

Модель триптофана ( Trp/W по коду tgg )

Модель тирозина ( Tyr/Y по коду tac )

Модель валина ( Val/V по коду gtc )